 2025, Vol. 17
2025, Vol. 17



2. 浙江大学环境与资源学院,浙江杭州 310058;
3. 国家林业和草原局华东调查规划院,浙江杭州 310019;
4. 中国科学院生态环境研究中心区域与城市生态安全全国重点实验室,北京 100085
2. College of Environmental and Resource Sciences, Zhejiang University, Hangzhou 310058, China;
3. East China Academy of Inventory and Planning, National Forestry and Grassland Administration, Hangzhou 310019, China;
4. State Key Laboratory of Regional and Urban Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
2024年全球平均气温比工业化前平均水平高出约1.55℃,是首个全球平均气温比工业化前水平高出1.5℃以上的年份,温室气体排放水平也再创纪录[1],共同应对气候变化已成为时代共识。森林碳汇是陆地生态系统中最大的碳库,而造林可以直接增加森林面积和生物量中储存的有机碳量,最终增强有机碳汇,因而植树造林工程成为许多国家重要的缓解气候变化的自然解决方案[2]。按储存介质分,在森林固定二氧化碳的过程中,从大气中吸收的碳主要储存在生物量(44%)和土壤有机质(45%)中,其余存在于枯枝落叶(6%)和枯木(4%)中[3]。其中土壤碳储量的变化相对来说较为缓慢,一般会随土地利用性质的变化而变化[4],枯枝落叶和枯木碳储量占比较小,而森林生物量碳储量占比较大,且其变化受到森林的生长和植被的动态变化影响,具有较大的固碳潜力[5]。
森林生物量碳储量的估算对于理解生态系统碳循环、应对气候变化具有关键意义,是评估造林等项目缓解气候变化效果的基础。国内外学者对此开展了大量研究,形成了多种评估方法及数据源应用体系。国外对森林生物量碳储量的研究开始于19世纪,德国林学家Ebermayer对乔木树干生物量的研究为后续工作奠定了基础[6, 7]。此后,欧洲、美国、日本等国家和地区相继展开测定与调查。国内研究起步于21世纪初[8-12],在国家、省市级尺度均取得显著成果。按照数据源的不同可以将目前常用的森林生物量碳储量估算方法分为基于森林清查数据和基于遥感数据的方法。
基于森林资源调查监测数据的方法是最经典的森林生态系统碳计量方法,森林资源调查监测数据也被认为是最可靠的数据来源[5]。该方法是通过设立固定样点及大规模的野外实地调查获取的一类清查样地数据,或者结合遥感判读和野外验证核实的二类调查图斑数据[13, 14],利用实测值或生物量转换因子法、平均生物量法、换算因子连续函数法、生物量回归方程法等模型先对森林生态系统各组成部分生物量进行测定估算,再通过碳转化系数将其转化为森林碳储量。方精云等[15]利用中国50年的国家森林资源清查数据,估算了1949—1998年森林碳储量的变化情况。李海奎和雷渊才[16]结合森林资源连续清查资料估算中国森林植被生物量和碳储量。近年来,国内外也有诸多学者对不同的生物量和碳储量估计方法进行了对比分析:李海奎等[17]比较了IPCC的通用方法、方精云的转换因子连续函数法和生物量回归模型法,认为生物量回归模型法的精度最高。Gao等[18]对4种碳储量评估方法进行了对比,认为直接采用碳储量回归模型效果最好。曾伟生等[12]使用三种方法对生物量和碳储量进行估算,认为基于林木水平的生物量模型法准确性最高,基于林分蓄积量的生物量转换因子法可改进可变参数提高适应性,基于蓄积量的换算因子连续函数法不适用于中小尺度应用[12]。利用森林清查数据进行的森林碳储量估算,虽然准确度较高,但需要投入大量人力和物力,且在固定样本点上进行连续调查的样地清查法全局性较差,而图斑数据在一类清查和二类调查数据的耦合衔接上还存在缺陷[13],在实际应用中仍具有一定的局限性。
基于遥感数据的方法一般结合了模型模拟法和遥感估算法。模型模拟法是基于植被生理过程,整合影响植被生态过程的环境因子与植被自身的生理特征,通过建立一定的数学关系模型,对森林生态系统生产力、生物量和碳储量进行估算研究的方法[19]。但许多模型基于静态或经验统计,带有较多假设,可能会在复杂的生态系统中存在许多不确定性。因此结合模型和利用遥感技术,能够更灵活地捕捉不同森林区域的碳储量变化,为大尺度碳循环过程提供更加全面和精确的评估与支持[20]。遥感估算法利用光学遥感、合成孔径雷达(Synthetic Aperture Radar, SAR)、激光雷达(Light Detection and Ranging, LiDAR)及多源融合数据[21],提取植被指数、地形因子、气象因子,与实测数据计算的碳储量进行拟合,建立碳储量与遥感数据的估算模型[22],实现森林生态系统碳储量的动态监测从样地到区域、全国甚至是全球尺度的转换。目前国内外学者在不同的空间尺度上基于不同类型的光学遥感影像数据,进行了众多森林碳储量及生物量的反演估算研究。如舒洋等[23]以Landsat-8遥感影像建立模型,张加龙和胥辉[24]利用遥感构建高山松地上生物量估测参数模型,梅晓丹[25]利用Biome-BGC模型估算了东北森林生态系统年碳通量及空间分布,王智等[26]采用CASA(Carnegie-Ames-Stanford Approach)模型对杭州市森林碳储量进行估算。碳储量估算软件模型也被大量开发,如InVEST、FORCCHN、CO2FIX、CITYgreen和i-Tree模型。
近年来,我国在立木生物量回归模型建立方面取得了大量成果,并颁布实施了一系列树种的立木生物量模型及碳计量参数标准[27]。同时,利用遥感数据进行自上而下的森林碳储量不同时空尺度估算的研究越来越多。以往研究表明,基于森林资源连续清查的样地和样木数据对生物量碳储量的估算利用了立木水平的生物量模型法,精度最高[12],但其工作量大、成本高,且难以实现在大尺度上对森林碳储量进行动态监测。基于遥感数据的估算尽管时空灵活性和可得性更强,但作为自上而下获取的数据源,其估算结果的准确性是否可以被接受仍需要在不同尺度上加以验证[20]。而图斑数据虽然耦合了一类清查、二类调查数据和遥感数据,解决了样地离散的主要问题,但其同样存在工作量大、成本高、更新不及时及监测精度得不到保障的问题[14]。当前我国生态环境管理正从粗放式治理向精细化调控转型,亟须科学、系统、动态评估林业工程的碳汇效益以支撑政策优化。但基于图斑数据、遥感数据的森林碳储量估算结果与基于样地数据的方法是否存在显著差异,以及差异的主要来源等缺乏区域尺度上的研究,可能导致管理决策偏差。基于以上现状与问题,本研究以福建省沿海防护林体系建设工程实施区为研究区域,尝试利用不同数据源估算森林碳储量。具体研究内容包括:①基于森林资源清查的样地、二类调查的图斑数据,和遥感反演的森林地上生物量产品及遥感解译的土地利用类型精细分类数据,估算森林碳密度和碳储量及其变化,分析其时空格局;②以样地结果为参考值,对比不同数据源的估算结果,分析其估算差异。研究将讨论产生估算差异的可能原因及不同数据源在碳储量估算中的优势与局限性,为沿海防护林工程的精准化管理和碳汇交易机制设计提供支撑。
1 研究区概况及数据来源 1.1 研究区概况本研究选取福建省33个沿海县(市、区)为研究区域(23.57°N~27.44°N,116.92°E~120.72°E,图 1)。研究区域位于中国东南沿海,属南亚热带和中亚热带湿润季风气候,年均温度及降水分别为20℃和1483 mm,地区海拔高度的范围为1~1791 m,陆地海岸线总长3752 km,主要的森林类型为常绿针叶林(48.89万hm2,44.73%)和混交林(30.77万hm2,28.15%),其余类型有常绿阔叶林(28.33万hm2,25.92%)和落叶阔叶林(1.31万hm2,1.20%),还有极少落叶针叶林。1988年福建省按照原国家林业部的统一部署,开始实施沿海防护林体系建设工程,旨在通过人工造林、封山育林、改造低效林等手段于沿海地区建立起一个以人工森林植被为主体的多林种、多树种、多功能、多效益的综合防护林体系。2022年,工程区土地总面积301.69万hm2,其中林地176.25万hm2,占比58% 左右。《福建省沿海防护林体系建设工程规划(2016—2025年)》提出进一步增加沿海地区森林面积和森林蓄积,提高森林覆盖率,是增加森林碳汇、积极应对全球气候变暖的必然选择[28]。

|
图 1 研究区域概况 |
本文选取三种数据源类型,分别为样地、图斑和遥感(表 1)。森林资源连续清查中的样地调查是以掌握宏观森林资源现状与动态为目的,以省(自治区、直辖市)为单位,利用固定样地为主进行定期复查的森林资源调查方法,是全国森林资源与生态状况综合监测体系的重要组成部分。每个省份都按照本省(区、市)安排,以五年的周期进行长期调查。而图斑数据也并非逐年更新,最早的完整记录为2014年,最新的一期数据为2022年。遥感数据相对灵活,能逐年获取。综合三类数据可获取的、可重叠分析的年份,我们选取了2014年和2022年的图斑和遥感数据,并选取了2013年的样地数据与2014年图斑和遥感数据对比(为方便描述,后续统一表述为2014年)。所用样地数据来自第八次全国森林资源清查福建省样地调查。样地调查是以样地为单元,采用地面调查与遥感判读相结合的方法,调查更新样地(样方)、样木因子,获取各类林草湿资源储量、质量、结构及其动态变化信息的过程[14]。每个样地面积为1亩(约667m2,即25.82 m×25.82 m)。样地数据是最准确的森林资源调查数据,通过定期对固定样地进行重复调查,可以准确监测森林资源在一定时间跨度内的动态变化。但样地调查成本较高,且数据更新周期长,时效性较差[29];同时作为抽样调查,它的空间连贯性差,无法体现区域尺度森林资源的空间格局。所用图斑数据来自福建省2014年和2022年二类调查数据。图斑监测是以小斑为单元,采用遥感判读和地面核实相结合的方法,监测图斑变化,更新图斑属性,获取各类林草湿资源面积、分布及其动态变化信息的过程[14]。相比于样地,多源数据融合的图斑数据能从宏观角度对大面积森林资源进行整体监测,全面覆盖区域;但对于一些微观层面的信息,如单株树木的胸径、树高、蓄积量等,图斑监测数据的精度较低。所用遥感数据来自公开数据库,包括森林地上生物量数据和土地覆盖数据,分辨率为30 m,时间为2014年和2022年,其中由于森林地上生物量数据未更新至2022年,沿用2021年。遥感是森林资源监测的重要数据源,其时空分辨率灵活性强,可实现动态监测;但其数据精度受多种因素影响,不确定性较强。
| 表 1 数据来源与说明 |
本研究对森林碳储量的估算均基于生物量法和平均含碳系数法。先基于样地、图斑、遥感数据的不同特征,选取不同生物量直接或间接估算方法计算2014年研究区域内林地的单位面积森林生物量和碳密度;再结合图斑和遥感数据的森林面积获得2014年和2022年的森林碳储量,然后计算两期碳储量的变化量为森林碳汇量;接着分析福建省沿海防护林工程实施区域内碳密度、碳储量的时空格局;并对比样地、图斑、遥感数据源下2014年的碳密度,以及图斑、遥感数据源下碳储量的估算结果差距;最后分析讨论差距的可能来源(图 2)。

|
图 2 技术路线 |
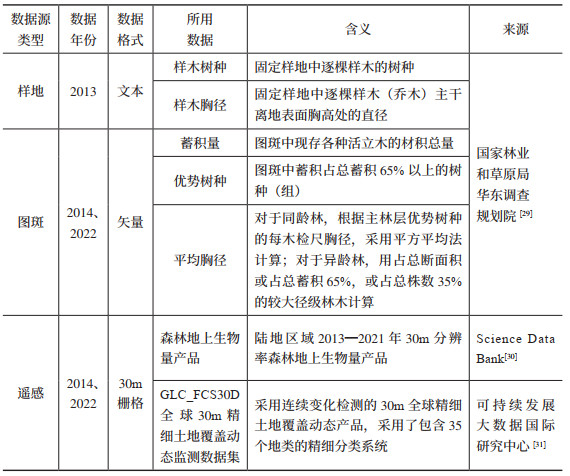
对于样地数据,采用生物量模型法和平均含碳系数法。首先为保证可比性筛选活立木蓄积大于零的林地样地;再基于样地中的样木树种确定立木生物量一元模型及碳计量参数,结合样木胸径计算出每个森林样地的地上、地下生物量,所用参数来源于公开文献与林业标准[16, 27, 33, 34],其中属于《主要树种立木生物量模型与碳计量参数》中21类主要树种的利用对应的基于胸径的生物量一元模型计算,其余树种按照通用模型[35]计算;最后根据含碳系数(Carbon Factor, CF)将生物量转换为碳储量。其中,含碳系数参考现有研究成果[16, 27],如果树种尚未测定含碳系数,则使用近似树种含碳系数或该地区树种的平均值代替[14, 20]。所用样地林木所属树种(组)及其平均胸径等数据来源于国家森林资源连续清查,森林有效样地为277个。碳密度的具体计算公式见式(1)和式(2):
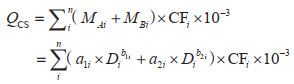
|
(1) |
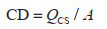
|
(2) |
其中,QCS是该样地当期所有样木地上和地下生物量总碳储量(Mg);MAi和MBi分别是该样地第i株样木的森林地上和地下生物量(kg);n是该样地的样木总株数;a1i、b1i、a2i、b2i是该样地第i株样木对应树种(组)立木一元生物量模型的参数;Di是该样地第i株样木的胸径(m);CFi是该样地第i棵树的平均含碳系数;CD是该样地的森林生物量碳密度(Mg/hm2);A是该样地面积(hm2),每块样地均为0.0667 hm2。
在计算完样地碳密度后,使用ArcGIS Pro的地统计分析模块进行协同克里金空间插值反演,获取整个研究区域森林的平均碳密度,以统一碳密度对比的空间尺度。克里金方法是地统计学中最常用的空间插值的方法,以变异函数为基础,可在一定区域内对某一变量未知位置进行线性最优无偏估计。协同克里金(cokriging)是一种基于克里金的空间插值技术,它通过考虑多个变量之间的空间相关性来优化预测[36, 37]。与传统的克里金方法相比,协同克里金不仅利用主变量(主要研究对象)的自相关性,还结合了其他辅助变量与主变量之间的互相关性,从而提高预测精度[38, 39]。其核心在于建立变量之间的交叉协方差函数,通过联合建模来提高预测的准确性。已有研究表明,协同克里金法适合于碳密度空间插值[36, 38, 40-43]。首先对包括海拔、坡度、土壤厚度、腐殖质厚度、植被覆盖度、林龄、平均胸径、平均树高、郁闭度、降水量、气温、NDVI在内的12个潜在协变量与2014年的碳密度分别进行相关性分析,选择相关性显著较高的3个变量(林龄、平均树高和郁闭度)作为协变量。其次,在森林样地中随机选取70% 用来进行半变异模型建模,称为训练集;剩余30% 用于精度验证,称作验证集。接着,为取得更好的估算效果,本研究利用ArcGIS Pro的地统计分析模块对训练集进行11种变异模型的计算和分析以选取最优模型,根据变异函数的理论和评价方法,得到最优变异函数的拟合结果,并采用标准平均值、标准均方根预测误差和块金比值3个指标对模型进行评价。一般来说最优模型的交叉验证的标准平均值的绝对值接近于0,标准化均方根误差接近于1,块金比值即模型所得块金值与基台值的比值表示空间变异性程度,分3个级别:0~<25%、25%~<75%、>75%,分别表示空间自相关程度为高等、中等和微弱。空间自相关程度越高,说明引起空间变异的主要是结构性因素,相反则主要是随机因素,只有在变量具备中等及以上的空间自相关性时,才可进行地统计空间插值[43]。根据表 2结果可知,不同半变异函数的拟合稳健性良好。另外,又通过验证集对所有模型进行了二次验证,在插值反演后的碳密度数据中提取与验证集样点相同位置的碳密度数据,进行精度检验与相关性分析,预测与真值的拟合线接近1 ∶ 1线被认为反映了更优的预测精度。综合训练模型和验证集验证获取的评价指标,最终选取K-Bessel模型来拟合。最后通过林斑图层与插值反演图层进行叠加与裁剪,从而得到2014年研究区域森林的碳密度空间分布格局图,并获取全域森林平均碳密度,作为来自样地数据源的对比依据。
| 表 2 协同克里金不同半变异函数评价指标 |

对于图斑数据,采用基于蓄积量的生物量转换因子法和平均含碳系数法。首先根据地类筛选出图斑中的林地;然后根据每个林地图斑的优势树种确定生物量转换因子(Biomass Conversion Factor, BCF)和根茎比(Root-to-Shoot Ratio, RSR),其中属于《主要树种立木生物量模型与碳计量参数》中21类主要树种的利用基于胸径的BCF、RSR一元模型计算[27],其余根据树种所属森林类型按照《中国森林生态系统生物量及其分配研究》中不同森林类型的平均BCF和RSR确定[33];CF取值方法与样地一致;最终计算得到每块林地图斑的碳密度和碳储量。但由于前后期图斑不固定,边界和森林面积均不一致,图斑数据的固碳速率将在转化为栅格后计算,仅表示该像元前后期碳密度的年均变化量,而未考虑其是否均为林地。相关公式如下[12, 44]:
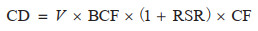
|
(3) |
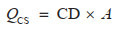
|
(4) |
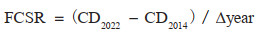
|
(5) |
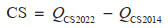
|
(6) |
其中,CD是该图斑的森林碳密度(Mg/hm2);V是该图斑当期单位面积活立木蓄积量(m3/hm2);BCF是该样地的平均生物量转换因子(Mg/m3);RSR是该图斑优势树种(组)的平均根茎比;CF是该图斑优势树种(组)的平均含碳系数。QCS是该林地图斑当期地上和地下生物量总碳储量(Mg);A是该图斑面积(hm2)。FCSR是森林生物量固碳速率[Mg/(hm2· a)];CD2022和CD2014分别为图斑转化成的栅格2022年和2014年的碳密度;Δ year取8年。CS是森林生物量碳汇量(Mg);QCS2022和QCS2014分别为一定区域内林地图斑2022年和2014年的碳储量总量。
2.3 基于遥感数据的计算方法对于遥感数据,直接基于森林地上生物量数据产品采用平均含碳系数法计算。首先获取研究区域内2014年和2021年森林地上生物量数据产品;然后利用GLC_FCS30D全球30 m精细土地覆盖动态监测数据2014年和2022年两期,选取开放常绿阔叶林、密闭常绿阔叶林、开放落叶阔叶林、密闭落叶阔叶林、开放常绿针叶林、密闭常绿针叶林、开放落叶针叶林、密闭落叶针叶林、开放混交林、密闭混交林10类森林类型,不同森林类型RSR和CF取福建省主要树种的平均值[16, 34];最终计算得到碳密度和碳储量。但同图斑数据,固碳速率仅表示该像元前后期碳密度的年均变化量。碳密度计算方法见式(7),碳储量、固碳速率和碳汇量公式同式(4)~式(6):
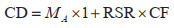
|
(7) |
其中,CD是该栅格的森林碳密度(Mg/hm2);MA是单位面积森林地上生物量(Mg/hm2);RSR是该森林类型的平均根茎比;CF是该森林类型的平均含碳系数。
3 结果与分析 3.1 森林碳密度和碳储量时空格局分析2014—2022年,总体上基于图斑数据的森林碳密度和碳储量估算结果在像元和县(市、区)尺度上均呈上升趋势,空间上呈现高值区集中在中部和南部的特征(图 3)。2014—2022年图斑数据估算的平均碳密度从31.29 Mg/hm2上升至38.20 Mg/hm2。相比于2014年,2022年碳密度在25 Mg/hm2以上的像元占比从49% 上升至70%,高值区分布在中部和南部地区。27% 的像元在8年间呈负碳汇趋势,其中部分可能是由于土地利用类型变为非林地,部分可能是森林类型及结构质量的变化导致碳密度下降。2014—2022年图斑数据估算的碳储量从35.33 Tg上升至44.93 Tg,在县(市、区)尺度上看,大多数沿海县的碳储量呈增加趋势,2014年只有仙游县的碳储量超过了3 Tg,2022年福安市、仙游县、漳浦县、诏安县均超过3 Tg,且只有四县(12%)的森林碳汇量为负,福安市、仙游县和罗源县在八年间产生了1 Tg以上的碳汇。

|
图 3 基于图斑数据的森林生物量碳密度、碳储量和碳汇量时空格局 |
基于遥感数据的森林碳密度和碳储量总体上在像元和县(市、区)尺度上均呈上升趋势,空间上呈现近海区域低、内陆山区高的特征(图 4)。相比于2014年,2022年森林碳密度整体增加(50.48~53.14 Mg/hm2),且碳密度在45 Mg/hm2以上的像元占比从56% 上升至73%。近海区域的森林碳密度较低,碳密度高值区主要分布在远离海岸的山区。35% 的像元呈负碳汇趋势,同样部分可能是源于土地利用类型的变化。2014—2022年遥感数据估算的碳储量从60.53 Tg上升至65.84 Tg,在县(市、区)尺度上,福安市、蕉城区、仙游县两期碳储量均高于5 Tg。2022年有85% 的县(市、区)产生了显著的森林碳汇,特别是福安市、南安市、霞浦县和福清县的森林在八年间产生了0.5 Tg以上的碳汇。

|
图 4 基于遥感数据的森林生物量碳密度、碳储量和碳汇量时空格局 |
比较2014年样地、图斑、遥感数据的碳密度估算结果,呈现图斑 < 样地 < 遥感的趋势,且不同结果之间均存在显著性差异(图 5)。根据福建省沿海县森林资源清查样地数据计算并插值得到的2014年森林碳密度约为33.34 Mg/hm2。基于立木水平的生物量模型法充分利用了森林资源清查中林木的胸径、树种等数据,通过实地样本拟合的模型参数来保证精度[45],被认为是精度最高的生物量及碳储量估算数据源[12, 17]。因此我们将使用样地的碳密度作为参考值,发现图斑数据估算的2014年碳密度为31.29 Mg/hm2,比样地低6.64%。图斑计算过程中涉及的BCF和RSR计算方法与样地类似,CF确定方法一致,因此推断偏低的碳密度可能是图斑数据测算的单位面积蓄积量或平均胸径偏低导致的。而遥感数据估算的碳密度比样地高51.41%,比图斑高61.34%,2022年的碳密度也比图斑高约39.12%。与样地使用立木水平的生物量模型相比,遥感数据直接利用了地上生物量图层,该数据集的交叉验证结果表明,与实地调查结果相比拟合结果总体偏高[40],因此差异大多来自于此。与图斑相比,遥感计算过程中涉及的RSR计算方法与图斑一致,差异来源可能来自地上生物量密度,经过统计分析,再次证明遥感数据源下偏高的碳密度可能与偏高的地上生物量密度数据有关,2014年和2022年遥感的平均地上生物量密度分别为82.73 Mg/hm2和91.10 Mg/hm2,分别高于图斑地上生物量密度(49.67 Mg/hm2和60.75 Mg/hm2)约66.57% 和49.95%。遥感数据源相对样地和图斑的偏高估算结果充分说明遥感来源的生物量数据还需要立足更精细的遥感纹理特征,结合更多的实地调查结果,针对森林碳储量的估算目标进一步提高模拟精度。

|
图 5 不同数据源碳密度估算结果对比 注:***P<0.001,表示两数据源均值比较存在显著差异。 |
比较2014年和2022年图斑、遥感数据的碳储量和碳汇量估算结果,发现图斑估算得到的两期碳储量较低,但两期间的碳汇量较高。如图 6a所示,图斑数据估算得到的福建省沿海防护林工程建设区域2014年和2022年的总碳储量分别为35.33 Tg和44.93 Tg,年碳汇量为1.20 Tg/a。而遥感估算的碳储量分别为60.53 Tg和65.84 Tg,分别比图斑结果高71.34%和46.55%,年碳汇量为0.66 Tg/a,比图斑结果低44.65%。碳储量的估计涉及碳密度和森林面积,图斑和遥感对碳储量的估计值差异一方面来自遥感估算的碳密度偏高,另一方面可能是对森林类型与面积的识别差异导致的。因此,我们在下节中进一步分析图斑和遥感数据对不同林分的识别与碳密度估算差异的影响。

|
图 6 不同森林类型的碳储量和面积对比 |
在研究森林碳储量和碳汇量的过程中,尺度效应是一个不可忽视的因素,本研究不仅关注了像元尺度和县(市、区)尺度的结果,还进一步拓展到林分尺度进行分析。基于图斑和遥感数据能统一分到的最细层次统计了不同森林类型的碳密度、碳储量和碳汇量结果。如图 7所示,总的来说,两种数据源密闭森林的碳密度均比开放森林高,这是由于随着覆盖度的提高,林分结构与质量整体有所提升,单位面积的碳蓄积更高。图斑结果显示2014年的密闭常绿阔叶林(50.70 Mg/hm2)和2022年的密闭落叶针叶林碳密度最高(62.10 Mg/hm2),两年的开放落叶阔叶林(7.53 Mg/hm2和9.35 Mg/hm2)碳密度均最低。遥感结果显示,两年的密闭常绿针叶林的碳密度最高(63.15 Mg/hm2和68.12 Mg/hm2),开放常绿针叶林最低(43.50 Mg/hm2和43.88 Mg/hm2)。经过t检验,图斑和遥感数据下不同森林类型的碳密度均有显著差异,除了密闭常绿阔叶林之外,图斑估算的其他类型的森林碳密度都低于遥感估算结果;且除了开放和密闭常绿阔叶林、密闭落叶阔叶林,其余森林类型两期估算得的碳密度差距都较大(大于一倍)。经分析,这与图斑估算的生物量密度和遥感使用的生物量密度的差距是一致的,而与计算过程中涉及的BCF和RSR的取值关系较小。

|
图 7 不同森林类型的碳密度估算结果对比 注:***表示P<0.001,表示两数据源均值比较存在显著差异。 |
对于碳储量和碳汇量,图斑和遥感数据估算的密闭常绿针叶林和密闭常绿阔叶林碳储量均最高(图 6a),图斑估算的密闭混交林碳汇量最高,而遥感估算的密闭常绿针叶林的碳汇量最高。虽然两期遥感识别的森林总面积仅比图斑高7% 左右,但最终遥感数据源估算得到的碳储量却分别比图斑高71.34%和46.55%。这部分差距除了与遥感估算得到的较高碳密度有关之外,还可能与两数据源识别的不同森林类型的面积差距过大有关(图 6b),特别是遥感数据对密闭常绿阔叶林等碳密度高的森林面积的识别(分别为106.27万hm2和103.23万hm2)显著多于图斑识别的面积(22.05万hm2和21.37万hm2),导致遥感计算得到的碳储量会更高。此外,遥感数据对高碳密度林分面积的识别偏差(如密闭混交林漏判)可能导致碳汇潜力被低估,建议在管理中结合地面核查优化遥感分类算法,提升监测精度。
4 讨论在全球气候变化的大背景下,森林作为重要碳汇,对减缓温室气体排放、维持生态平衡意义重大,特别是在受气候变暖威胁更大的沿海地区。森林碳储量估算关乎林业工程的实施调整、碳排放交易等应对气候变化机制的运行。但森林生态系统复杂、区域森林类型多样、测量方法有差异,这些都导致碳储量估算存在误差[46],从而影响决策科学性,不利于林业体系工程的价值实现。因此本研究基于三种碳储量估算常用数据源的特征,使用生物量法和平均含碳系数法,以福建省沿海防护林工程建设区域为例进行实证对比分析,对森林碳储量的科学评估具有指导作用。
首先,从时间格局上看,福建省沿海区域的森林碳密度和碳储量呈增加趋势,这不仅在样地尺度的碳密度升高上得到验证,图斑数据和遥感数据的碳密度、碳储量结果也反映相同的趋势。碳密度的升高可能是由于该地区日照和水源充足,水热条件适宜林木生长,森林的质量得到提升(包括森林的年龄、胸径、树高等),因此尽管2014—2022年区域内森林面积有所下降,但碳密度的提升却使得碳储量得到提升[47, 48]。20世纪80年代以来,福建省在沿海地区实施的沿海防护林体系建设工程等林业生态工程加大了区域内的造林、保护和抚育力度,有效抑制了森林的损失,调整了林龄结构,这些都有助于增加森林的碳储量[49]。这表明沿海防护林在降低大气CO2浓度方面具有重要作用,沿海防护林体系的建设具有重要的生态效益,未来需在政策层面保障工程延续性,制定长期生态规划,一方面培育已有林地,优化林种结构与林龄配置,提高林分质量,另一方面通过造林增加林地面积,提高碳汇普惠性,并将碳汇增量纳入地方政府生态绩效考核体系。另外,实施精准抚育,对成熟林分开展疏伐、修枝等抚育作业,促进林木胸径与树高增长;对幼龄林实施补植补造,优先选择速生、高碳汇树种。
其次,从空间格局上看,靠近海岸线的区域森林分布较稀疏,碳密度和碳储量也相对较低。一方面这可能是由于沿海地区城镇化程度和人为干扰程度更高,对自然生态系统的开发及人类活动的增强导致林地的直接减少与森林的退化;另一方面碳密度在内陆—沿海间显示显著的差异与Zhao等[50]的研究结论类似,该研究发现离岸距离对中国沿海红树林保护区的总初级生产力变化的影响较大。沿海环境压力较大,近海地区的海风会通过机械和干旱胁迫影响植被的发育,也可能运输盐雾,影响森林冠层、茎密度和林分面积调节水分和土壤水分有效性,从而影响地上生物量[51]。有研究还发现在靠近海岸线的地区土壤有机层薄、营养不足会导致植被营养缺乏症的增加[52],影响沿海植被的正常生长。为协调生态保护与经济发展,建议加强林分管理,如在近海防护林带优先推广耐盐碱树种(如木麻黄、红树林),并设置缓冲区减少人为干扰,同时通过土壤改良(如添加有机质、施用缓释肥)提升立地质量;对于受近海恶劣环境影响较小的内陆地区,可以重点保护、增植碳密度高的树种(如合适的常绿阔叶林和针叶林),并实施补植补造以优化林分结构,适当采取人工促进更新和增汇的措施,并通过生态补偿机制激励社区参与防护林养护。
对比不同数据源的估算结果,发现2014年福建省沿海地区的森林碳密度呈现图斑(31.29 Mg/hm2)<样地(33.34 Mg/hm2)<遥感(50.48 Mg/hm2)的趋势,2022年碳密度同样呈现图斑(38.20 Mg/hm2)<遥感(53.14 Mg/hm2)的趋势,在不同森林类型尺度上大致也呈同样的趋势。对于图斑数据来说,其与样地数据的结果差距较小,这体现了国家森林资源清查数据的一致性。细究其估算使用的数据,图斑数据的碳密度和碳储量利用基于蓄积量的生物量转换因子法计算,主要与该图斑每公顷蓄积量、优势树种和平均胸径有关。相较于样地数据源下立木水平的生物量模型法,该方法得到的碳密度偏低,与曾伟生等[12]的研究结论类似。由于图斑是对森林斑块平均状况的估计,很有可能不能充分统计森林实际的蓄积量,基于蓄积量的生物量转换因子法会将蓄积量估计的误差相应地带到生物量的估计中,如果蓄积量估计值偏低,其生物量及后续的碳密度估计值也会偏低。其不确定性还有部分来自不在21类主要树种中的其他树种的BCF和RSR参数直接使用了全国平均值,这可能并未体现福建省本土林木的特征。对于遥感数据来说,其碳密度主要基于地上生物量密度数据(R2值在0.611和0.682,与验证点相比偏高)和根据土地利用精细分类数据(总体精度为82.5%)赋值的BCF和RSR平均值,这两类数据都是结合卫星遥感和地面调查拟合而成的模型结果[47],囿于遥感的影像分辨率、数据处理技术及地形效应等多重因素的局限性,其各自存在一定误差,叠加耦合的过程又不可避免地引入了新的误差来源[46],导致其可能无法监测到森林实际的细微变化,因此虽然遥感数据能极大提升大尺度测量的效率与覆盖范围,但其仅能反映森林植被的整体生长状况,缺乏对其他因子的考虑,其对森林碳密度的估算存在一定误差。
对比分析图斑和遥感两数据源下森林碳储量及不同森林类型的碳储量,图斑总碳储量明显低于遥感,但碳汇量高于遥感,而不同类型森林的碳储量结果差异较大。图斑和遥感数据估算得到的碳储量和碳汇量的差异一方面来自上述提到的两数据源下估算得到的碳密度的差异,另一方面来自对不同类型森林面积的识别。相对来说,由于图斑监测结合了遥感判读和大规模的地面核实,我们认为图斑数据对不同森林类型的面积识别是较为可靠的。而遥感数据对不同森林类型识别的误差可能源于多方面,如光谱特征相似致不同类型易混淆、分类精度与土地覆盖异质性呈负相关[53]、时序影像空间分辨率较低或时间相隔较长且不连续[54]等,均可能导致识别精度有限[55]。因此出现图斑和遥感数据对于不同类型森林面积的识别差距大,叠加碳密度的误差,导致不同森林类型碳储量的差距,从而导致总碳储量的差异。
总而言之,三种数据源在估算森林碳储量过程中有利有弊。尽管已明确样地数据源准确度更高,但在实际操作过程中仍存在测量误差和模型误差,如样地代表性不足、测量工具精度不高、调查人员的主观误差等测量误差[56],以及模型样本的不全面、参数的不确定性、非主要树种的通用模型准确度低等模型误差[46]。另外,更为关键的是囿于人力物力成本,仅能以固定抽样的形式对区域内的森林样地进行定期调查,导致其缺乏空间连贯性,要想获得区域内的森林总体数量和质量,还需要结合遥感等技术。遥感技术作为现代森林碳储量估算的重要手段,具有成本低、时空连续性好等优点,而且可以克服单一使用森林清查数据所带来的人为统计的主观不确定性[57],且本研究运用的土地利用精细分类已是国际上时间跨度最长、分类体系最精细的全球30 m土地覆盖遥感产品。相对来说,图斑是样地数据与遥感数据耦合之后的外推,图斑尺度森林碳储量能够较好反映森林资源的植被特征,能综合实地调查获得的林分蓄积量,并结合遥感判读和地面核查而实现较为准确的估算。但同样地,在衔接一类样地清查和二类图斑调查的过程中必然存在偏差[13],这是由于在将核实样地信息扩展到图斑时,可能未充分考虑森林生态系统的空间异质性,即同一图斑内的林木,其实际树种、蓄积本身就存在差异,而在图斑数据的生成过程中可能往往进行平均化处理。另外,森林资源图斑调查数据的更新速度较慢,而且数据的获取较为困难,也具有一定的局限性[58]。
在未来森林碳汇估算与管理工作中,可从以下方面深入探索:一是强化样地数据与遥感数据的融合。样地数据精确但覆盖范围有限,遥感数据能宏观快速获取大面积信息却存在局部细节不足。因此建议基于样地数据、图斑数据和深度学习算法,建立一套更精细的生态系统类型分类数据,然后再结合遥感影像,反演整个区域的各类生态系统调节服务(包括碳固定、水源涵养、土壤保持、空气净化等)为生态补偿、生态产品交易等提供可靠的数据基础。二是拓展生物量模型,提升其适用性和准确性。现有模型普适性有限,未来应广泛收集不同气候带、生态系统和经营模式下的森林生物量数据,运用多元统计、机器学习等方法优化,考虑引入土壤质地、地形坡度、气候因子等环境变量,拓展不同树种的专门模型,满足不同研究和应用需求[59]。三是持续在不同空间尺度检验不同数据的估算差异。森林生态系统空间异质性明显,小尺度聚焦单个林分或地块研究测量方法差异,中尺度针对特定区域分析数据源优缺点,大尺度以国家或洲际范围探讨估算方法的宏观适用性。通过系统检验,为研究者提供参考,助力其精准选择合适估算方法,提升估算工作的科学性和有效性。
5 结论系统分析不同数据源对森林碳储量估算的误差,识别可能的误差来源,择优获取更准确的估算结果,既有助于深化对森林碳循环机制的理解,也能为应对气候变化策略的规划与管理提供量化支撑,特别是在受气候变暖威胁更大的沿海地区。本研究基于森林资源清查样地数据、图斑数据和遥感数据,分别计算了福建省沿海防护林工程建设区域内森林碳密度、碳储量及碳汇量,除了进一步明确了福建省沿海防护林体系建设工程实施区域内碳密度、碳储量的时空格局,还通过对比分析发现:①以样地数据为参考值,图斑数据计算得到的碳密度偏低,遥感数据偏高,分析差距的可能原因是图斑数据对蓄积量的测量存在误差、遥感生物量数据准确度不佳;2014年和2022年图斑数据估算得到的总碳储量均低于遥感数据,但碳汇总量高于遥感,差距可能来源于森林面积的识别偏差。②从不同森林类型角度,遥感估算的大部分类型的森林碳密度都高于图斑结果,差距可能来自遥感生物量数据的偏差;两个数据源估算的碳储量存在显著差异,通过分析再次印证差距来自对不同森林类型的识别。总的来说,样地数据能最准确地估算碳密度,图斑数据能相对准确地识别森林类型和面积,未来需要继续加强样地数据和遥感数据的多元融合,提高生物量模型的准确度和适用性,完善多源数据协同监测体系,建立碳汇动态评估平台,从而实现不同尺度森林碳储量准确、连续、便捷的评估,并强化政策衔接,将碳汇核算结果纳入生态补偿标准制定与生态绩效考核。
| [1] |
World Meteorological Organization (WMO). WMO confirms 2024 as warmest year on record at about 1.55℃ above pre-industrial level[EB/OL]. (2025-01-10)[2025-02-18]. https://wmo.int/news/media-centre/wmo-confirms-2024-warmestyear-record-about-155degc-above-pre-industrial-level.
|
| [2] |
LU N, TIAN H Q, FU B J, et al. Biophysical and economic constraints on China's natural climate solutions[J]. Nature climate change, 2022, 12(9): 847-853. DOI:10.1038/s41558-022-01432-3 |
| [3] |
Food and Agriculture Organization of the United Nations (FAO). Global Forest Resources Assessment 2020: Main Report[M]. Rome: FAO, 2020.
|
| [4] |
侯瑞萍, 夏朝宗, 陈健, 等. 长江经济带林地和其他生物质碳储量及碳汇量研究[J]. 生态学报, 2022, 42(23): 9483-9498. |
| [5] |
刘迎春, 高显连, 付超, 等. 基于森林资源清查数据估算中国森林生物量固碳潜力[J]. 生态学报, 2019, 39(11): 4002-4010. |
| [6] |
王新闯, 齐光, 于大炮, 等. 吉林省森林生态系统的碳储量、碳密度及其分布[J]. 应用生态学报, 2011, 22(8): 2013-2020. |
| [7] |
郭娇宇. 内蒙古东部主要人工林碳计量参数及生物量模型研究[D]. 呼和浩特: 内蒙古农业大学, 2021.
|
| [8] |
刘国华, 傅伯杰, 方精云. 中国森林碳动态及其对全球碳平衡的贡献[J]. 生态学报, 2000, 20(5): 733-740. DOI:10.3321/j.issn:1000-0933.2000.05.004 |
| [9] |
王效科, 冯宗炜, 欧阳志云. 中国森林生态系统的植物碳储量和碳密度研究[J]. 应用生态学报, 2001, 12(1): 13-16. DOI:10.3321/j.issn:1001-9332.2001.01.003 |
| [10] |
ZENG W S. Development of monitoring and assessment of forest biomass and carbon storage in China[J]. Forest ecosystems, 2014, 1(1): 20. DOI:10.1186/s40663-014-0020-5 |
| [11] |
FANG J Y, CHEN A P, PENG C H, et al. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 2001, 292(5525): 2320-2322. DOI:10.1126/science.1058629 |
| [12] |
曾伟生, 陈新云, 蒲莹, 等. 基于国家森林资源清查数据的不同生物量和碳储量估计方法的对比分析[J]. 林业科学研究, 2018, 31(1): 66-71. |
| [13] |
曾伟生. 森林资源调查监测中的数据耦合方法研究[J]. 林业资源管理, 2022(2): 61-66. |
| [14] |
国家林业和草原局. 2022年全国森林、草原、湿地调查监测技术规程[EB/OL]. (2022-04-30). [2023-08-17]. https://ghhzrzy.tj.gov.cn/zwgk_143/tzgg/202206/W020220606558259191558.pdf.
|
| [15] |
方精云, 陈安平. 中国森林植被碳库的动态变化及其意义[J]. 植物学报, 2001, 43(9): 967-973. DOI:10.3321/j.issn:1672-9072.2001.09.014 |
| [16] |
李海奎, 雷渊才. 中国森林植被生物量和碳储量评估[M]. 北京: 中国林业出版社, 2010.
|
| [17] |
李海奎, 赵鹏祥, 雷渊才, 等. 基于森林清查资料的乔木林生物量估算方法的比较[J]. 林业科学, 2012, 48(5): 44-52. DOI:10.3969/j.issn.1006-2505.2012.05.016 |
| [18] |
GAO H L, DONG L H, LI F R, et al. Evaluation of four methods for predicting carbon stocks of Korean pine plantations in Heilongjiang Province, China[J]. PLoS one, 2015, 10(12): e0145017. DOI:10.1371/journal.pone.0145017 |
| [19] |
赵振宇. 基于森林资源清查和遥感数据的青浦区森林碳储量估算及空间分布研究[D]. 上海: 上海师范大学, 2023.
|
| [20] |
胡海波, 刘佳璇, 丁冬霞, 等. 森林固碳计量方法研究综述[J]. 中南林业科技大学学报, 2024, 44(11): 58-69. |
| [21] |
章范怡. 基于多源遥感数据的森林地上生物量及碳储量估算方法研究[D]. 南京: 东南大学, 2023.
|
| [22] |
冯国平. 基于多源数据的驻马店建成区绿地碳储量估算研究[D]. 郑州: 河南农业大学, 2024.
|
| [23] |
舒洋, 周梅, 赵鹏武, 等. 大兴安岭南段华北落叶松人工林碳储量及分配特征研究[J]. 生态环境学报, 2016, 25(10): 1604-1611. |
| [24] |
张加龙, 胥辉. 基于遥感的高山松连清固定样地地上生物量估测模型构建[J]. 北京林业大学学报, 2020, 42(7): 1-11. |
| [25] |
梅晓丹. Biome-BGC模型参数优化及东北森林碳通量估算研究[D]. 哈尔滨: 东北林业大学, 2017.
|
| [26] |
王智. 基于CASA模型的杭州市森林碳储量时空变化及影响因子研究[D]. 杭州: 浙江农林大学, 2021.
|
| [27] |
国家林业和草原局. GB/T 43648-2024主要树种立木生物量模型与碳计量参数[S]. 北京: 中国标准出版社, 2024.
|
| [28] |
福建省林业局. 福建省林业局关于印发《福建省沿海防护林体系建设工程规划(2016-2025年)》的通知[R]. 福州: 福建省林业局, 2019.
|
| [29] |
福建省林业厅. 第九次全国森林资源清查福建省清查操作细则[R]. 福州: 福建省林业局, 2018.
|
| [30] |
YAN S J, HE G J, ZHANG X M. Forest aboveground biomass products in China, 2013-2021[DB/OL]. Science Data Bank, 2023. [2025-04-14]. https://doi.org/10.57760/sciencedb.07122.
|
| [31] |
刘良云. GLC_FCS30D: 1985-2022年全球30米精细土地覆盖动态监测数据集[DB/OL]. 可持续发展大数据国际研究中心, 2023. [2025-04-19]. https://data.casearth.cn/thematic/glc_fcs30/314.
|
| [32] |
邓成, 梁志斌. 国内外森林资源调查对比分析[J]. 林业资源管理, 2012(5): 12-17. DOI:10.3969/j.issn.1002-6622.2012.05.003 |
| [33] |
罗云建, 王效科, 张小全, 等. 中国森林生态系统生物量及其分配研究[M]. 北京: 中国林业出版社, 2013.
|
| [34] |
北京林业大学. LY/T 2988-2018森林生态系统碳储量计量指南[S]. 北京: 中国标准出版社, 2019.
|
| [35] |
ZENG W S, TANG S Z. A new general allometric biomass model[J]. Nature Precedings, 2011, 6: 1. |
| [36] |
贺鹏, 张会儒, 雷相东, 等. 基于地统计学的森林地上生物量估计[J]. 林业科学, 2013, 49(5): 101-109. DOI:10.3969/j.issn.1672-8246.2013.05.018 |
| [37] |
WEN W, WANG Y F, YANG L, et al. Mapping soil organic carbon using auxiliary environmental covariates in a typical watershed in the Loess Plateau of China: a comparative study based on three Kriging methods and a soil land inference model (SoLIM)[J]. Environmental earth sciences, 2015, 73(1): 239-251. DOI:10.1007/s12665-014-3518-9 |
| [38] |
王璟睿. 结合陆地卫星与森林资源连续清查数据的森林碳密度估算研究[D]. 南京: 南京林业大学, 2017.
|
| [39] |
LONG J, LIU Y L, XING S H, et al. Optimal interpolation methods for farmland soil organic matter in various landforms of a complex topography[J]. Ecological indicators, 2020, 110: 105926. DOI:10.1016/j.ecolind.2019.105926 |
| [40] |
杜雪, 王景弟, 白彦锋, 等. 基于克里金插值法的湖南省慈利县森林碳储量专题图研究[J]. 西北林学院学报, 2022, 37(1): 198-204. DOI:10.3969/j.issn.1001-7461.2022.01.29 |
| [41] |
邱新彩, 郑冬梅, 王海宾, 等. 结合地统计学与Landsat 8影像的乔木林地上碳储量估算[J]. 中南林业科技大学学报, 2020, 40(11): 138-146. |
| [42] |
李妙宇, 上官周平, 邓蕾. 黄土高原地区生态系统碳储量空间分布及其影响因素[J]. 生态学报, 2021, 41(17): 6786-6799. |
| [43] |
张峰, 杜群, 葛宏立, 等. 基于地统计学和CFI样地的浙江省森林碳空间分布研究[J]. 生态学报, 2012, 32(16): 5275-5286. |
| [44] |
国家发展和改革委员会, 国家统计局. 生态产品总值核算规范[M]. 北京: 人民出版社, 2022.
|
| [45] |
华伟平, 邱宇, 徐波, 等. 基于生长生物量模型法的福建森林碳汇估算研究[J]. 西南林业大学学报, 2014, 34(6): 35-43. |
| [46] |
普发全. 森林碳储量估算中的误差分析与改进方法[J]. 中国林业产业, 2024(9): 22-23. |
| [47] |
ZHANG X M, HE G J, YAN S J, et al. Forest status assessment in China with SDG indicators based on high-resolution satellite data[J]. International journal of digital earth, 2023, 16(1): 1008-1021. DOI:10.1080/17538947.2023.2190625 |
| [48] |
黄从德, 张健, 杨万勤, 等. 四川森林植被碳储量的时空变化[J]. 应用生态学报, 2007, 18(12): 2687-2692. |
| [49] |
冯源, 肖文发, 朱建华, 等. 造林对区域森林生态系统碳储量和固碳速率的影响[J]. 生态与农村环境学报, 2020, 36(3): 281-290. |
| [50] |
ZHAO D M, ZHANG Y H, WANG J J, et al. Spatiotemporal dynamics and geo-environmental factors influencing mangrove gross primary productivity during 2000-2020 in Gaoqiao Mangrove Reserve, China[J]. Forest ecosystems, 2023, 10: 100137. DOI:10.1016/j.fecs.2023.100137 |
| [51] |
MEDINA E, CUEVAS E, MOLINA S, et al. Structural variability and species diversity of a dwarf Caribbean dry forest[J]. Caribbean journal of science, 2010, 46(2-3): 203-215. DOI:10.18475/cjos.v46i2.a9 |
| [52] |
LIU B Y, WANG Y, CHEN Z H, et al. Coastal dwarf forest, the new phytosociological unit or the enlargement of current syntaxa?[J]. Ecosystem health and sustainability, 2023, 9: 0064. DOI:10.34133/ehs.0064 |
| [53] |
ZHANG X, LIU L Y, CHEN X D, et al. GLC_FCS30: global land-cover product with fine classification system at 30 m using time-series Landsat imagery[J]. Earth system science data, 2021, 13(6): 2753-2776. DOI:10.5194/essd-13-2753-2021 |
| [54] |
徐凯健. 基于高分卫星多尺度时空特性的区域森林优势树种遥感识别[D]. 南京: 南京大学, 2019.
|
| [55] |
王长委, 胡月明, 沈德才, 等. 基于CBERS数据的亚热带森林地上碳储量估算[J]. 林业科学, 2014, 50(1): 88-96. DOI:10.3969/j.issn.1006-1126.2014.01.018 |
| [56] |
邓蕾, 上官周平. 基于森林资源清查资料的森林碳储量计量方法[J]. 水土保持通报, 2011, 31(6): 143-147. |
| [57] |
丛俊霞, 郑晓, 朱教君, 等. 沙地樟子松天然林地上碳储量估算及其空间分布特征[J]. 生态学杂志, 2017, 36(11): 2997-3007. |
| [58] |
黄从红, 杨军, 张文娟. 森林资源二类调查数据在生态系统服务评估模型InVEST中的应用[J]. 林业资源管理, 2014(5): 126-131. |
| [59] |
张聪, 刘琪, 李海奎, 等. 我国尺度兼容和树种分类的材积源森林碳储量模型[J]. 林业科学, 2025, 61(1): 57-69. |